
 Cuando era niño me advirtieron que las agujetas se producían por culpa del lactato, de sus cristales puntiagudos, y que debía tomar mucho azúcar para evitarlas. Desde hace algunos años esta idea está desacreditada. Pero el lactato sigue asumiendo el papel protagonista en la fatiga. Se considera al lactato una sustancia venenosa que produce acidez muscular y que impide que el organismo pueda rendir a pleno rendimiento. Estamos ante un nuevo error, que me gustaría reparar aportando información científica sobre el papel real que desempeña el lactato en el rendimiento deportivo y en la fatiga, una sustancia esencial para el correcto funcionamiento de nuestro organismo y a la que resulta necesario reintegrarle su inocencia.
Cuando era niño me advirtieron que las agujetas se producían por culpa del lactato, de sus cristales puntiagudos, y que debía tomar mucho azúcar para evitarlas. Desde hace algunos años esta idea está desacreditada. Pero el lactato sigue asumiendo el papel protagonista en la fatiga. Se considera al lactato una sustancia venenosa que produce acidez muscular y que impide que el organismo pueda rendir a pleno rendimiento. Estamos ante un nuevo error, que me gustaría reparar aportando información científica sobre el papel real que desempeña el lactato en el rendimiento deportivo y en la fatiga, una sustancia esencial para el correcto funcionamiento de nuestro organismo y a la que resulta necesario reintegrarle su inocencia.
La respiración humana y por tanto, nuestra forma de acceder a la energía, es un calco inverso de la fotosíntesis. La principal molécula energética del reino animal es la glucosa, que sólo las plantas pueden producir gracias a la energía solar. A través de la glucosa el ser humano acaba utilizando para su metabolismo la energía solar que las plantas procesaron. Si ellas tomaron dióxido de carbono y agua y lo mezclaron para producir glucosa, nosotros la separaremos gracias al oxígeno, para producir la energía que necesitamos para vivir.
GLUCOSA + 6 O2 = 6 CO2 + 6 H2O + ENERGÍA
La oxidación de un mol de glucosa (180 gramos) genera unas 689 kcal (unas 4 kcal por cada gramo de glucosa), una pequeña parte de las cuales se utilizan para formar moléculas de ATP (la moneda energética del ser humano, a razón de 32 moléculas de ATP por cada molécula de glucosa), y el resto, se disipará en forma de calor (66%). Tan sólo 233 kcal (34%) podrán ser utilizadas como energía libre, por el ser humano, por cada mol de glucosa oxidada.
Esta reacción oxidativa tan simple, el organismo humano no la realiza de una vez, sino a través de gran número de reacciones químicas intermedias (en cascada), con gran presencia de enzimas, y fundamentalmente desarrolladas en el citoplasma celular y sobre todo dentro de las mitocondrias. A través de esta cascada de reacciones intermedias el ser humano va extrayendo poco a poco la energía contenida originalmente en la glucosa, produciéndose una gran variedad de productos intermedios.
Este largo proceso de conversión de la glucosa en energía (y anhídrido carbónico y agua) se puede desarrollar, además, siguiendo dos caminos complementarios, las llamadas vías aeróbicas y anaeróbicas. Que se siga una u otra no depende de la presencia o ausencia de oxígeno, sino del ritmo al que se requiere generar energía y de la mayor o menor concentración, en los músculos solicitados, tanto de mitocondrias, como de las correspondientes enzimas de cada una de estas vías alternativas, que finalmente concluyen en el resultado reflejado en la ecuación anterior. El organismo nunca opta entre una u otra vía, sino que siempre utiliza las dos a la vez, eso sí, en diferente proporción según los músculos implicados y la intensidad del esfuerzo desarrollado.
Conviene resaltar que el oxígeno lo utiliza tanto la vía aeróbica como la anaeróbica, pero que en ambos casos el oxígeno sólo participa al final de todas las reacciones, y que hasta ese momento final en el que el oxígeno recoge los electrones contenidos en los iones hidrógeno (H+) generados, todas las reacciones químicas se realizan en ausencia de oxígeno, tanto en la vía aeróbica como en la anaeróbica. Por estas razones, creo que resulta más apropiado denominar a estas vías alternativas y complementarias, como de lácticas (anaeróbica) y alácticas (aeróbica), también por la razón que a continuación explicaremos.
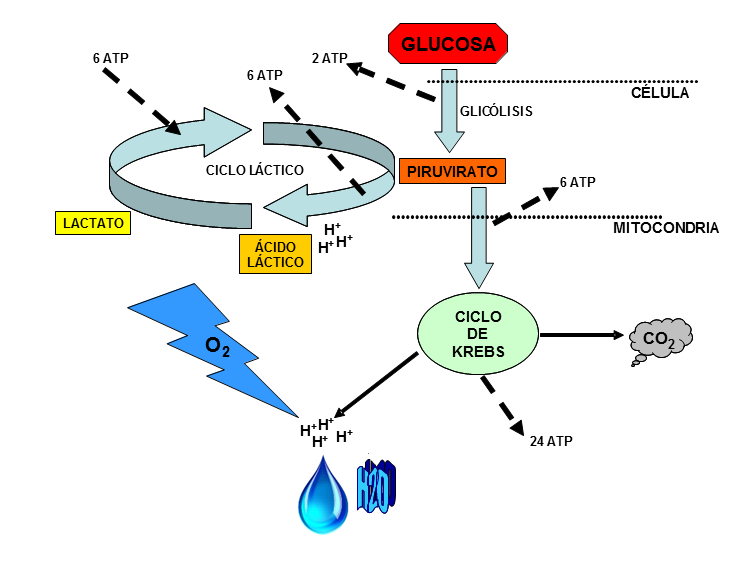
Ambas vías comparten unas reacciones comunes en el tramo inicial de oxidación de la glucosa, las llamadas reacciones de glicólisis, que transforman la glucosa en energía (2 moléculas de ATP) y en un compuesto denominado piruvirato, que contiene el resto de la energía no extraída y originalmente contenida en la glucosa. La glicólisis se desarrolla en el citoplasma, fuera de las mitocondrias, sin presencia de oxígeno, y los biólogos consideran que esta reacción química es un arcaísmo que todavía utilizan, por ejemplo, reptiles, peces y mamíferos marinos para procurarse energía rápidamente.
Una vez producido, el piruvirato penetra en la mitocondria, y es aquí donde empieza la vía láctica o aláctica, según el piruvirato se transforme en lactato o penetre en el llamado ciclo del ácido cítrico (ciclo de Krebs). A la vía láctica también se la denomina rápida, ya que a través de menos reacciones ofrece energía con más celeridad, aunque en menor cantidad. Y a la aeróbica, lenta, porque obtiene más energía tras un proceso más largo. Pero téngase en cuenta que en un músculo el piruvirato que se va generando tras la glicólisis, en unos puntos se estará transformando en lactato, y en otros será asimilado por la respiración aeróbica (alaáctica), dos vías que necesariamente hay que considerar como complementarias, y no como alternativas o excluyentes.
Conviene también aclarar y resaltar el hecho de que la llamada vía rápida o láctica comparte con la lenta o aláctica, el tramo final de reacciones químicas. Es decir, que el organismo humano lo que hace con la vía láctica (rápida) es obtener poca energía, pero muy rápido, para responder a una demanda muscular intensa, pero que el resto de la energía no extraída en primera instancia de la glucosa, se extraerá finalmente en la vía aláctica previa conversión del lactato otra vez en piruvirato. Por ello, la vía láctica celular no aporta energía neta, ya que la energía liberada al transformarse el piruvirato en lactato, tiene que ser utilizada de nuevo para convertirlo más tarde otra vez en piruvirato. Lo útil de este proceso neutro a nivel energético, es la liberación rápida de energía, que más tarde, cuando el organismo reduzca el ritmo de ejercicio, será empleada nuevamente para comenzar el ciclo aláctico. Los períodos de relajación entre series intensas poseen esta finalidad. El ciclo láctico, por tanto, posee la única utilidad de ayudar a coordinar necesidad energética con producción, pero el resultado final siempre será el mismo. Ambas vías, por lo que se ve, se dan siempre juntas, y lo que cambia, según los músculos, la intensidad y las características fisiológicas del atleta, es la particular coordinación y peso relativo de cada vía en la producción de energía.
En resumen, el ser humano extraerá 32 moléculas de ATP desde una de glucosa, en un solo proceso lento (la vía aláctica) y directo, o a través de un desvío muy rápido que pasa por la generación de lactato y pequeñas dosis de energía (2 ATP), pero que finalmente converge en la vía aláctica, con una ganancia energética neta similar en ambos procesos, pero con distintas velocidades de obtención.
Como se aprecia en la figura, casi toda la energía se produce en el ciclo del ácido cítrico (24 ATP), y el oxígeno sólo aparece al final del proceso, en el mismo punto, ya sea a través de la vía láctica o aláctica. Luego el que se opte por el desvío rápido (láctica) o por la vía directa más lenta (aláctica) dependerá no de la falta de oxígeno (que nunca se produce) sino del ritmo al que los músculos demandan energía y del cóctel enzimático que posea cada individuo. Pero aclaremos este último punto un poco más.
Un individuo que posea una proporción elevada de fibras musculares de contracción rápida, y por tanto, que tenga, en principio, un complejo enzimático adaptado a la generación de gran potencia muscular, sus mitocondrias elegirán predominantemente la vía láctica para generar energía. Puede decirse, que incluso a ritmos relativamente bajos, generará elevadas dosis de lactato respecto a un individuo dotado de un mayor porcentaje de fibras de contracción lenta. El que el individuo rápido y potente pueda hacerse también resistente dependerá de su capacidad para introducir lactato (previamente convertido en piruvirato) dentro del ciclo del ácido cítrico (vía lenta o aláctica), o sea, de que el entrenamiento sea capaz de incrementarle las enzimas y mitocondrias que participan en las reacciones de esta última vía metabólica. El que el individuo lento y resistente se haga también potente, dependerá de su capacidad para incrementar su metabolismo láctico en conjunción, sobre todo, con el aumento de las enzimas y las mitocondrias que servirán para quemar ese incremento de lactato.
Pero las dos afirmaciones anteriores conviene interpretarlas a la luz del siguiente razonamiento. Que cada reacción individual de la vía aláctica sea más lenta, no quiere decir que también lo sea la suma de todas las reacciones alácticas que se desarrollen en un determinado músculo. Si el músculo posee una concentración de mitocondrias y enzimas “alácticas” muy elevada, su funcionamiento en cascada podrá producir un flujo de energía también muy rápido. Por esta razón un corredor resistente dotado de un elevado porcentaje de fibras de contracción lenta puede correr a una velocidad de crucero también muy elevada, porque muchas mitocondrias lentas, debidamente coordinadas pueden generar un flujo constante y potente de energía. Sin embargo, esta elevada concentración no le dará la facultad de cambiar el ritmo con rapidez. Esto es lo que ocurre, por ejemplo, con el miocardio, que sólo tiene capacidad para usar la vía aláctica, y que posee una concentración elevadísima de mitocondrias. ¿Quién duda de su capacidad para generar altas dosis de energía? Pero su ritmo, en cambio, posee más inercias que las de un músculo de contracción rápida. Su capacidad tanto de incrementar su ritmo (respuesta) como de disminuirlo (recuperación) dependerá de la mejor o peor coordinación de sus mitocondrias a la hora de modular su flujo de producción de energía con las demandas de los músculos esqueléticos.
Estamos acostumbrados a considerar al oxígeno en su papel de activador de la combustión, y por tanto, solemos entender que el proceso de generación de energía desde la glucosa se produce también por combustión, como si la glucosa ardiera en el oxígeno. Pero resulta más apropiado interpretar el papel del oxígeno en la respiración humana en su extraordinaria capacidad para captar electrones (oxidar). Recordemos que la glucosa se formaba en ese laboratorio químico que son las hojas de las plantas, gracias a que la clorofila es capaz de extraer la energía del sol “robándole” los electrones con los que construirá la glucosa. Todo el proceso que hemos explicado de obtención de energía desde esta glucosa se realiza liberando esos electrones que la planta le quitó a la energía solar, en forma de iones hidrógeno (H+). Si no existiera un método de evacuación o de neutralización de esos iones, el ph de las células musculares se haría cada vez más ácido, imposibilitando las contracciones musculares, e incluso sus mínimas funciones vitales. Y aquí es precisamente cuando aparece el oxígeno, en el tramo final tanto de las vías lácticas y alácticas, para llevarse estos iones en forma de agua (O- + 2 H+ = H2O), y que expiramos junto con el CO2 formado previamente en el ciclo respiratorio.
El motor humano es muy distinto al de combustión de un coche, aunque el símil se utilice en muchas ocasiones. En ambos el oxígeno resulta esencial, pero por diferentes motivos. El automóvil necesita llevar consigo todo el combustible que va a quemar, los más de 50 litros del depósito los introducirá en los cilindros para recorrer, digamos 500 kilómetros. Si del mismo modo el ser humano debiera llevar todo el combustible consigo, tendría que acarrear más de 80 kilogramos de ATP para correr, por ejemplo, una maratón, cuando de hecho el cuerpo humano sólo almacena 85 gramos de ATP. Tan sólo acarreamos glucógeno y grasas, de las que consumiremos menos de un kilogramo. Nuestro metabolismo, en cambio, se parece más al de una pila eléctrica, ya que no son reacciones de combustión sino de tipo redox (reducción-oxidación) las que se producen a nivel celular para generar energía y movimiento: los electrones de la glucosa circulan como la corriente eléctrica, desde zonas de mayor potencial (glucosa) hasta el vapor de agua que sale de los pulmones. Y es en este camino en el que se interponen las moléculas de ATP, que transfieren esta energía del flujo de electrones a los procesos vitales que hacen posible el movimiento y la vida. Las células no pueden consumir ni calor ni electricidad, únicamente pueden utilizar la energía química contenida en los enlaces fosfóricos del ATP, que se erige así en el gran trasbordador energético del mundo celular. Lo asombroso de nuestro metabolismo reside en que existe muy poco ATP en las células, ya que el ATP según se produce se consume, tan sólo es un intermediario que no se acumula en ningún depósito, sino que como la moneda, circula a mayor o menor velocidad según el ritmo de la actividad económica o muscular. Y la adecuación de la oferta energética con la demanda muscular se logra gracias al acoplamiento de las dos vías metabólicas láctica (lenta) y aláctica (rápida), que en ambos casos generan una acidez (iones H+) que habrá que evacuar finalmente gracias a la enorme capacidad oxidativa del oxígeno.
En el desvío de la vía láctica ocurre que para acelerar el ritmo de obtención de energía a partir del piruvirato, las mitocondrias utilizan reacciones y enzimas diferentes a las de la respiración aláctica y generan un subproducto llamado ácido láctico, que atesora todavía mucha de la energía original de la molécula de glucosa y que tiene encomendada una doble misión: la primera, unirse a los iones H+, convertirse en lactato e impedir la acidez muscular, y en segundo lugar, ya como lactato, acabar convirtiéndose otra vez en piruvirato para culminar su proceso de oxidación y por tanto, de extracción de energía, en el que los H+ momentáneamente unidos al lactato ahora se liberan y se evacuan finalmente por medio del oxígeno. El lactato así producido lo consumen los propios músculos, y también el corazón, al que le llega por el fluido sanguíneo, ya que el miocardio consume mayoritariamente lactato cuando trabaja intensamente. Y el hígado, que transformará el lactato en glucógeno (ciclo de Cori) que almacenará o que enviará otra vez a los músculos. Incluso músculos ociosos o poco requeridos, transformarán su glucógeno en lactato para transferirlo vía sanguínea a los músculos más solicitados, ya sea directamente como lactato, o como glucosa tras pasar por el hígado (transbordador de lactato).
El lactato producido no provoca ningún efecto negativo en el funcionamiento muscular. Si el ejercicio se hace muy intenso y la velocidad de síntesis de lactato a partir del ácido láctico y de los iones H+ liberados, no fuera tan rápida, entonces el lactato no tendría capacidad para evacuar toda la acidez muscular y serían los iones H+ y no el lactato el responsable de la disminución del rendimiento. Ocurre que este fenómeno se inicia cuando ya la concentración de lactato en sangre es realmente elevada, y se tiende a identificar lactato con disminución de la capacidad física, pero en realidad ha sido la incapacidad del organismo para producir más lactato la causante de la acidosis muscular.
En este momento ocurren dos fenómenos de gran interés. En primer lugar, y como mecanismo defensivo, el organismo, con el objetivo de reducir la acidez, empieza a derivar carbonato sódico (suplemento que utilizan los atletas de media distancia) a los músculos con objeto de que reaccione con los iones H+ y así reducir la acidez (reacciones tampón o buffer). Esto provoca la liberación extra de CO2, que se une al anhídrido carbónico y al vapor de agua propios de la respiración celular, provocando que el atleta deba hiperventilar para sacar de los pulmones ese exceso de CO2. A este punto en que empieza el estrés respiratorio se le ha llamado en ocasiones el segundo umbral ventilatorio, pero no coincide exactamente, como suele afirmarse, con el umbral anaeróbico, ni con el umbral láctico.
En segundo lugar, y cuando la intensidad del ejercicio se relaja, la concentración de lactato seguirá creciendo inicialmente, en un intento de incrementar la evacuación de iones H+ vía oxidación del ácido láctico. Hasta que el consumo de lactato acabe superando al de producción y finalmente se reestablezca el equilibrio. Durante la recuperación, por tanto, el organismo consumirá oxígeno extra a media que utiliza lactato como combustible, con objeto de limpiar al organismo de los iones H+ liberados por el lactato al convertirse en piruvirato y entrar en el ciclo de Krebs. A este oxígeno suplementario que introducimos en el cuerpo cuando disminuimos el ejercicio y nos recuperamos se le ha denominado “déficit de oxígeno”, porque en cierta manera su inspiración compensaba el oxígeno que habíamos dejado de consumir cuando la intensidad del ejercicio nos obligó a utilizar la vía láctica (anaeróbica). Por tanto, la medición o estimación de este déficit que se compensa posteriormente al ejercicio se utiliza con cierta frecuencia para estimar su intensidad. Con este uso encontraremos que se le denomina EPOC (Excess post oxygen consumption), o exceso de consumo de oxígeno posterior al ejercicio.
En fin, que el lactato ni provoca agujetas, ni fatiga, que estamos ante una sustancia esencial para el correcto funcionamiento del cuerpo humano, y que ya es hora de que empecemos a nombrar a los procesos fisiológicos que ocurren durante el ejercicio con propiedad y corrección, en honor de la verdad y de la correcta planificación del entrenamiento.

Elogio del lactato by Rui Valdivia is licensed under a Creative Commons Reconocimiento-NoComercial 4.0 Internacional License.

{kind=link}
Deja un comentario